Séminaire Dr Denise Arico
Colle moléculaire chez les plantes : LecRK-I.9 crée des connexions entre la membrane plasmique et la paroi cellulaire - Lundi 11 mai 2026 11h00, INRAE, Versailles
Lorsque les plantes sont exposées à un stress hyperosmotique intense, l’eau quitte la cellule, entraînant une plasmolyse, au cours de laquelle la membrane plasmique se détache partiellement de la paroi cellulaire. Ce détachement met en évidence des sites discrets où la membrane plasmique reste connectée à la paroi. Afin d’identifier les composants moléculaires à l’origine de cette liaison physique, nous avons réalisé un criblage ciblé de protéines candidates chez arabidopsis (Fig. 1). Parmi celles-ci, la surexpression de la kinase de type récepteur LecRK-I.9 a significativement augmenté la densité des connexions membrane plasmique–paroi dans les cellules de pavage chez Nicotiana benthamiana.
Nous avons constaté que le domaine lectine extracellulaire de LecRK-I.9 est nécessaire à l’établissement de ces connexions. En conditions hyperosmotiques, LecRK-I.9 forme des amas immobiles présentant une distribution asymétrique à la surface des cellules de l’épiderme dans différents organes d’Arabidopsis. La délétion du domaine lectine augmente la mobilité de ces amas et abolit cette asymétrie, ce qui indique que ce domaine régule la distribution spatiale de LecRK-I.9 à la membrane plasmique (Fig. 2).
Nous cherchons actuellement à déterminer si le domaine lectine se lie à des microdomaines spécifiques de la paroi et à évaluer si la surexpression de LecRK-I.9 module la déformation de la paroi lors d’un stress hyperosmotique. Nous évaluons également la tolérance de lignées surexprimant LecRK-I.9 à des conditions de faible potentiel hydrique et de sécheresse, qui mettent à l’épreuve l’intégrité du continuum membrane plasmique–paroi.
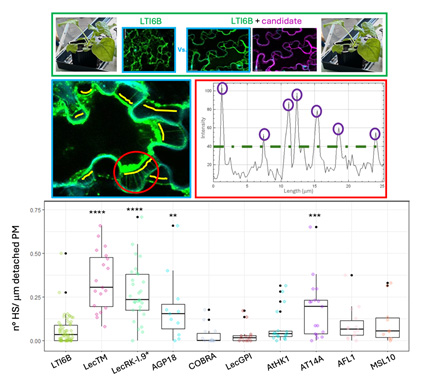 Figure 1. Criblage des protéines favorisant les connexions entre la membrane plasmique et la paroi cellulaire. Transformation transitoire de N. benthamiana suivie d’une plasmolyse des cellules de pavage avec du glycérol à 10 %, et quantification de la densité des filaments hechtiens marqués avec LTI6B-mCitrine. Boîte à moustaches montrant la densité des filaments hechtiens (FH), exprimée en nombre de FH par µm de membrane plasmique (MP) détachée, pour chaque candidat testé comparé au contrôle LTI6B. La significativité statistique a été déterminée à l’aide du test de Kruskal–Wallis suivi du test post hoc de Dunn avec correction de Benjamini–Hochberg. **** valeurs de p < 10⁻⁵, *** valeurs de p < 0,0002, ** valeurs de p < 0,003.
Figure 1. Criblage des protéines favorisant les connexions entre la membrane plasmique et la paroi cellulaire. Transformation transitoire de N. benthamiana suivie d’une plasmolyse des cellules de pavage avec du glycérol à 10 %, et quantification de la densité des filaments hechtiens marqués avec LTI6B-mCitrine. Boîte à moustaches montrant la densité des filaments hechtiens (FH), exprimée en nombre de FH par µm de membrane plasmique (MP) détachée, pour chaque candidat testé comparé au contrôle LTI6B. La significativité statistique a été déterminée à l’aide du test de Kruskal–Wallis suivi du test post hoc de Dunn avec correction de Benjamini–Hochberg. **** valeurs de p < 10⁻⁵, *** valeurs de p < 0,0002, ** valeurs de p < 0,003.
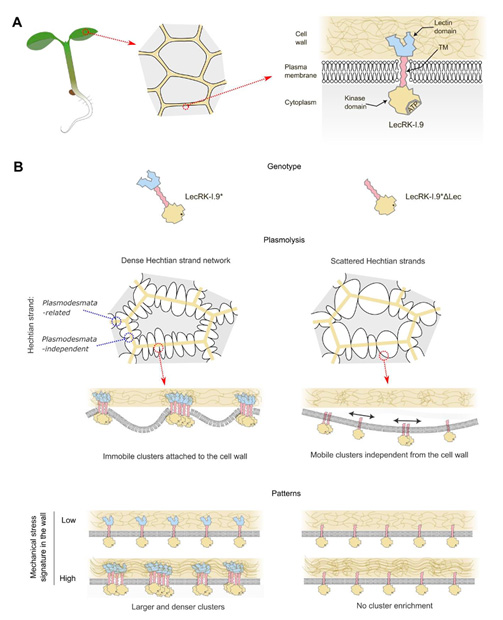 Figure 2. Modèle : LecRK-I.9 agit comme une colle moléculaire, ancrant les filaments hechtiens et reflétant la signature de contrainte mécanique dans la paroi. A. Description et localisation de LecRK-I.9. B. Résumé des résultats avec LecRK-I.9* et LecRK-I.9ΔLec. LecRK-I.9 : kinase de type récepteur à lectine de type légumineuse I.9. * 5 substitutions d’alanine aux positions 350 aa – 354 aa au sein du motif de liaison à l’ATP. LecRK-I.9ΔLec : LecRK-I.9 dépourvue du domaine lectine (Δ244 aa aux positions 22 aa – 265 aa).
Figure 2. Modèle : LecRK-I.9 agit comme une colle moléculaire, ancrant les filaments hechtiens et reflétant la signature de contrainte mécanique dans la paroi. A. Description et localisation de LecRK-I.9. B. Résumé des résultats avec LecRK-I.9* et LecRK-I.9ΔLec. LecRK-I.9 : kinase de type récepteur à lectine de type légumineuse I.9. * 5 substitutions d’alanine aux positions 350 aa – 354 aa au sein du motif de liaison à l’ATP. LecRK-I.9ΔLec : LecRK-I.9 dépourvue du domaine lectine (Δ244 aa aux positions 22 aa – 265 aa).
Denise Arico, "Méchanotransduction et développement" MECANODEVO, RDP, ENS de Lyon
Invitation : Grégory Mouille, équipe "Adhésion Cellulaire et Communication Intercellulaire" ACCI
Séminaire relatif à la recherche développée à l’Institut Jean-Pierre Bourgin - Sciences du Végétal.
Nous avons constaté que le domaine lectine extracellulaire de LecRK-I.9 est nécessaire à l’établissement de ces connexions. En conditions hyperosmotiques, LecRK-I.9 forme des amas immobiles présentant une distribution asymétrique à la surface des cellules de l’épiderme dans différents organes d’Arabidopsis. La délétion du domaine lectine augmente la mobilité de ces amas et abolit cette asymétrie, ce qui indique que ce domaine régule la distribution spatiale de LecRK-I.9 à la membrane plasmique (Fig. 2).
Nous cherchons actuellement à déterminer si le domaine lectine se lie à des microdomaines spécifiques de la paroi et à évaluer si la surexpression de LecRK-I.9 module la déformation de la paroi lors d’un stress hyperosmotique. Nous évaluons également la tolérance de lignées surexprimant LecRK-I.9 à des conditions de faible potentiel hydrique et de sécheresse, qui mettent à l’épreuve l’intégrité du continuum membrane plasmique–paroi.
Denise Arico, "Méchanotransduction et développement" MECANODEVO, RDP, ENS de Lyon
Invitation : Grégory Mouille, équipe "Adhésion Cellulaire et Communication Intercellulaire" ACCI
Séminaire relatif à la recherche développée à l’Institut Jean-Pierre Bourgin - Sciences du Végétal.
Retour
