Un transport d'hexoses entre le cytosol et la vacuole est nécessaire au développement des vaisseaux du xylème dans la tige de l'inflorescence
Les sucres ont des rôles variés dans le développement et la croissance des plantes.
Les sucres représentent la principale source d'énergie pour la cellule végétale et ils jouent le rôle de « briques de base » pour la synthèse des polysaccharides de la paroi cellulaire. Ils sont également essentiels en tant que signaux cellulaires. En effet, ils possèdent une activité morphogénétique et agissent comme messagers primaires dans les voies de transduction du signal. Chez les plantes, les tissus vasculaires se composent du phloème, du procambium/cambium et du xylème. C’est un système élaboré responsable du transport de la plupart des composés biologiques dans toute la plante, y compris les sucres. Au niveau moléculaire, le développement du système vasculaire est régi par un réseau complexe de facteurs de transcription qui sont sous le contrôle de plusieurs signaux, notamment des hormones, des peptides et des microARN. Cependant, dans ce cadre bien organisé, une certaine plasticité est nécessaire pour s'adapter aux variations cellulaires en termes de disponibilité des nutriments (c'est-à-dire les sucres et les acides aminés). Il est donc logique que des modifications du métabolisme, du transport ou de la signalisation des sucres puissent entraîner de multiples défauts dans la croissance et le développement des plantes. Cependant, malgré ce rôle central, le rôle de la disponibilité des sucres dans le développement du système vasculaire en général et plus spécifiquement dans les tissus hétérotrophes tels que le cambium et le xylème est encore mal compris. Le transport latéral des sucres est particulièrement crucial pour la formation de la paroi cellulaire secondaire du xylème puisqu’ils sont des composés intermédiaires dans la synthèse des polysaccharides de la paroi cellulaire, ces derniers représentant 80 % de la paroi secondaire. Le xylème joue un rôle de puit pour les sucres qui doivent être importés des tissus environnants pour servir de source de carbone et d'énergie. Cette hypothèse est étayée par le fait que les perturbations du transport des sucres au niveau de la membrane plasmique des cellules vasculaires, par l'intermédiaire des transporteurs de sucre (SWEET) ou de sucre (SUC/SUT), affectent la composition de la paroi cellulaire secondaire du xylème tant chez le tremble (Populus tremuloides) que chez les tiges d'inflorescence d'arabidopsis (Arabidopsis thaliana).
Cependant, la question, de l'importance de la modification de la répartition des sucres dans les cellules vasculaires, reste ouverte.
La vacuole représente le principal organite de stockage de nombreux métabolites primaires et spécialisés, dont les sucres, dans la cellule.
C’est au niveau du tonoplaste, la membrane de la vacuole, que s’effectue les échanges de sucres requis pour l'ajustement dynamique de la quantité de sucre nécessaire aux voies métaboliques et de signalisation. Chez les plantes herbacées et ligneuses, peu de transporteurs de sucres ont été caractérisés fonctionnellement au niveau du tonoplaste, SWEET16, et SWEET17 sont parmi eux. SWEET16 et 17 ont été principalement caractérisés par les équipes CATS et SATURNE à l'IJPB qui développent des études sur l'allocation du carbone et de l'azote depuis plus de 10 ans. Les transporteurs SWEET16 et SWEET17 sont localisés dans les racines. De plus, il a été précédemment montré que le promoteur SWEET16 est actif dans les cellules du parenchyme du xylème, tandis que le promoteur SWEET17 est actif dans les cellules du parenchyme du xylème et dans les jeunes cellules du xylème de la tige de l'inflorescence d'arabidopsis. De plus, des niveaux élevés de transcrits SWEET17 ont été mesurés dans la tige d'inflorescence, par rapport aux autres organes, y compris les racines, après 7 à 8 semaines de croissance. SWEET16 et SWEET17 sont donc de bons candidats pour évaluer si le maintien de l'homéostasie des sucres entre le cytosol et la vacuole influence le développement du xylème chez arabidopsis.
Ce travail montre que les transporteurs de sucre SWEET16 et SWEET17 ont des rôles spécifiques et complémentaires au cours du développement du xylème.
En particulier, les échanges tonoplastiques de sucres, régulés par SWEET16, à l’interface procambium-xylème, sont importants pour la prolifération des cellules du xylème. En utilisant la spectroscopie infrarouge à transformée de Fourier (FTIR) et l'analyse de l'expression des gènes, ce travail montre également que SWEET16 et SWEET17 sont tous deux nécessaires au développement correct de la paroi cellulaire secondaire des cellules du xylème. Enfin, comme une accumulation de glucose et de fructose est observée dans la tige d'inflorescence du double mutant, cette étude suggère que le maintien de l'homéostasie des hexoses par l'action de SWEET16 et/ou SWEET17 est important à différents stades du développement du xylème.
Cette étude a permis d'enrichir le modèle du rôle des transporteurs SWEET au cours du développement du xylème dans les tiges d'inflorescence d'Arabidopsis.
Cependant, des recherches complémentaires sont nécessaires afin de mieux comprendre comment la cellule hiérarchise la distribution des sucres entre les différents processus. Répondre à cette problématique permettrait de mieux comprendre l'allocation des sucres dans la plante et son impact, entre autres, sur la composition des graines.
Modèle concernant le rôle des transporteurs SWEET pendant le développement du xylème dans les tiges d'arabidopsis
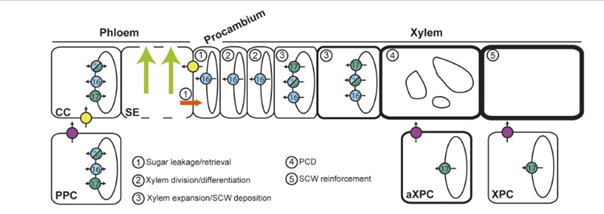
CC : cellules compagnes
Cercles fuchsia : SWEET11 et SWEET12
PCD : mort cellulaire programmée
PPC : cellules du parenchyme phloèmien
SCW : paroi cellulaire secondaire
SE : tubes criblés
XPC : cellules du parenchyme xylémien
Cercles jaunes : transporteur SUC2
Pour plus de détails, voir la publication
Retour

Agrandissement, lien
Legende : SWEET16 et SWEET17 sont exprimés dans les cellules du xylème tout au long du développement de la tige de l'inflorescence et forment des hétérodimères. A, C, et E. Analyse histochimique de l'activité GUS dans les lignées exprimant les protéines de fusion SWEET16-GUS sous l’action du promoteur natif de SWEET16 dans des sections prises à différentes positions dans la tige de l'inflorescence de plantes âgées de 7 semaines. B, D et F, analyse histochimique de l'activité GUS dans les lignées exprimant les protéines de fusion SWEET17-GUS sous l’action du promoteur natif de SWEET17 dans des sections prélevées à différentes positions dans la tige de l'inflorescence de plantes âgées de 7 semaines. Les sections ont été choisies dans une région de la tige où la croissance était encore rapide (A, B), dans une région de la tige où la croissance par élongation était terminée mais où l'épaississement de la paroi cellulaire secondaire était encore en cours (C, D), et au bas de la tige, une région qui correspond à une tige mature (E, F). Les flèches pointent les cellules présentant une coloration bleue issue de l’expression de GUS dans les cellules du xylème en développement, les têtes de flèches indiquent les cellules du parenchyme axial et les astérisques indiquent les cellules du parenchyme xylémien. La lignine est colorée en rose par du phloroglucinol. L'intensité de la couleur rose indique le niveau de lignification des vaisseaux du xylème.
ep, épiderme ; co, cortex ; iff, fibres interfasciculaires ; ph, phloème ; xy, xylème.
Fait marquant IJPB
Equipe "Carbone, Allocation, Transport, Signalisation"
CATS
Publication associée
Aubry, E, Beate Hoffmann, B, Françoise Vilaine, F, Françoise Gilard, F, Klemens PAW, 4 , Florence Guérard, F, Bertrand Gakière, B, Ekkehard Neuhaus, Bellini, C, Dinant, S, Le Hir, R. A vacuolar hexose transport is required for xylem development in the inflorescence stem. (2022) Plant Physiol. 188. https://doi.org/10.1093/plphys/kiab551