A vacuolar hexose transport is required for xylem vessels development in the inflorescence stem
Sugars play multiple and essential roles in plants
Plant sugars represent the main source of energy, are « building blocks » for the cell wall polysaccharides synthesis and play also a crucial role as signaling molecules. They arbor a morphogenetic activity and, act as primary messengers in signal transduction pathways. The plant vasculature, composed of phloem, procambium/cambium, and xylem, is an elaborate system responsible for the transport of most biological compounds throughout the plant including sugars. At the molecular level, vasculature development is governed by a complex network of transcription factors that are under the control of several signals, including hormones, peptides, and microRNAs. However, within this well-organized framework, a certain plasticity is required to adjust to cellular variations in terms of the availability of nutrients (i.e. sugars and amino acids). It is therefore logical that modifications of sugar metabolism, transport or signaling can lead to multiple defects in plant growth and development. However, despite this central role, the role of sugar availability in the development of the vascular system in general and more specifically in heterotrophic tissues such as cambium and xylem is still elusive. Lateral transport is especially crucial for xylem secondary cell wall formation, since sugars are intermediate compounds in the synthesis of the cell wall polysaccharides, which represent 80% of the secondary cell wall. The xylem tissue thus represents a strong sink for sugars that must be imported from surrounding tissues to serve as the source of carbon and energy. This is supported by the fact that perturbations in sugar transport at the plasma membrane of vascular cells, via SUGAR WILL EVENTUALLY BE EXPORTED TRANSPORTERS (SWEET) or SUCROSE TRANSPORTERS (SUC/SUT), affect the composition of the xylem secondary cell wall both in aspen (Populus tremuloides) and in Arabidopsis (Arabidopsis thaliana) inflorescence stems.
However, the question remained open as to whether the modification of sugar partitioning within the vasculature cells is of importance.
The vacuole represents the main storage compartment for numerous primary and specialized metabolites including sugars in the cell. At the level of its membrane, the tonoplast where sugar exchanges between the vacuole and the cytosol occur are therefore required for dynamic adjustment of the quantity of sugar needed for metabolic and signaling pathways. In herbaceous and ligneous plants, few sugar transporters have been functionally characterized at the tonoplast, SWEET16, and SWEET17 are among them. SWEET16 and 17 have been mainly characterized by the CATS and SATURNE teams at IJPB who developed studies on carbon and nitrogen allocation for more than 10 years. SWEET16 and SWEET17 transporters are localized in the roots. In addition, it has been previously shown that the SWEET16 promoter is active in the xylem parenchyma cells, while the SWEET17 promoter is active in the xylem parenchyma cells and young xylem cells of the Arabidopsis inflorescence stem. Moreover, high levels of SWEET17 transcripts have been measured in the inflorescence stem, compared to other organs including roots, after 7 to 8 weeks of growth. SWEET16 and SWEET17 are therefore good candidates with which to assess whether the maintenance of sugar homeostasis between the cytosol and the vacuole influences xylem development in Arabidopsis.
This work shows that the sugar transporters SWEET16 and SWEET17 have specific and overlapping roles during xylem development.
In particular, tonoplastic sugar exchanges across the procambium-xylem boundary regulated by SWEET16 are important for xylem cell proliferation. By using infrared spectroscopy and gene expression analysis, this work also shows that both SWEET16 and SWEET17 are required for correct development of the secondary cell wall of xylem cells. Finally, since glucose and fructose accumulation is observed in the inflorescence stem of the double mutant, this study suggests that maintenance of hexose homeostasis through the action of SWEET16 and/or SWEET17 is important at different stages of xylem development.
This study allowed to enrich a model for the role of SWEET transporters during xylem development in Arabidopsis inflorescence stems.
Further work is required to understand how the cell is prioritizing the distribution of sugars among the different processes, it remains an open question. This would help to better understand sugar allocation in plant and how it impacts among others seed composition.
Model for the role of SWEET transporters during xylem development in Arabidopsis stems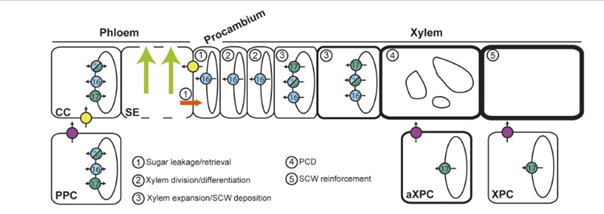
Model legend:
CC: companion cells
Fuchsia circles: SWEET11 and SWEET12
PCD: programmed cell death
PPC: phloem parenchyma cells
SCW: secondary cell wall
SE: sieve tubes
XPC: xylary parenchyma cells
Yellow circles: SUC2 transporter
16 / 17: SWEET 16 /17
For more details see the publication
Back

Full size, link
Legend: SWEET16 and SWEET17 are expressed in xylem cells throughout inflorescence stem development and form heterodimers. A, C, and E. Histochemical analysis of GUS activity in lines expressing SWEET16-GUS fusion proteins driven by the SWEET16 native promoter in sections taken at different positions in the inflorescence stem of 7-week-old plants. B, D, and F, Histochemical analysis of GUS activity in lines expressing SWEET17-GUS fusion proteins driven by the SWEET17 native promoter in sections taken at different positions in the inflorescence stem section of 7-week-old plants. Sections were taken in a stem region where the growth was still rapid (A, B, and insets), in a stem region where elongation growth had finished but where thickening of the secondary cell wall was still ongoing (C, D), and at the bottom of the stem, a regionn that corresponds to a mature stem (E, F, and insets). Arrows point to cells showing blue GUS staining in developing xylem cells and arrow heads point to axial parenchyma cells and asterisks indicate xylary parenchyma cells. Lignin is colored pink after phloroglucinol staining. The intensity of the pink color indicates the level of lignification of the xylary vessels.
ep, epidermis; co, cortex; iff, interfascicular fibers; ph, phloem; xy, xylem
Scientific Highlight of IJPB
"Carbon, Allocation, Transport and Signaling" team
CATS
Associated publication
Aubry, E, Beate Hoffmann, B, Françoise Vilaine, F, Françoise Gilard, F, Klemens PAW, 4 , Florence Guérard, F, Bertrand Gakière, B, Ekkehard Neuhaus, Bellini, C, Dinant, S, Le Hir, R. A vacuolar hexose transport is required for xylem development in the inflorescence stem. (2022) Plant Physiol. 188. https://doi.org/10.1093/plphys/kiab551